In the ever-evolving landscape of biomedical research, innovative tools play a pivotal role in unlocking new insights into complex diseases like cancer. Among these tools, cellular stains have emerged as indispensable assets, enabling scientists to visualise and interrogate cellular processes with unprecedented clarity and precision.
Join us as we embark on a journey through the latest advancements in cancer research, fueled by the ingenuity of cellular stains. Here, we selected five recent publications, showcasing the transformative impact of these stains on elucidating key molecular mechanisms, identifying novel therapeutic targets, and driving advancements in personalised medicine.
1. Targeting pancreatic cancer with genetically engineered membrane nanoparticles
Zhang et al. developed a bispecific single-chain variable fragment (scFv)-based nanoengager aimed at enhancing macrophage phagocytic activity by targeting tumour cells specifically. Through genetic engineering of cell membranes, the authors obtained bispecific scFv comprising anti-CD40 scFv (targeting macrophages) and anti-CLDN18.2 scFv (targeting tumour cells). These scFv were subsequently used to coat a PLGA core, resulting in nanoengagers that present both anti-CD40 scFv and anti-CLDN18.2 scFv. These nanoengagers significantly augmented immune responses, including heightened recognition and phagocytosis of tumour cells by macrophages, enhanced activation and antigen presentation, and elevated cytotoxic T lymphocyte activity. These combined advantages led to enhanced antitumour efficacy against highly aggressive “cold” pancreatic cancer. In summary, this study presents a versatile nanoengager design for immunotherapy, achieved through genetic engineering to incorporate antibody-anchored membranes.
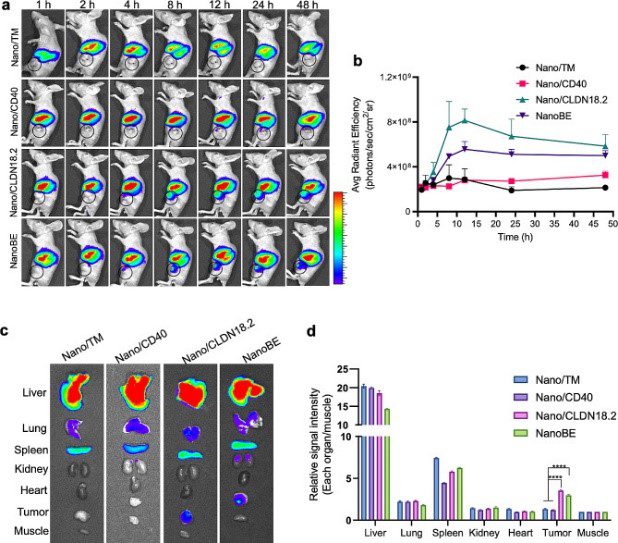
DiD and DiO, both lipophilic carbocyanine dyes, were used as lipophilic tracers to stain the membranes of KPC cells expressing anti-CD40 scFv and anti-CLDN18.2 scFv for the preparation of the nanoengagers. They were also utilised in co-culture experiments for the phagocytosis assay. Additionally, DiR (see figure below) was used to label the nanoengagers for near-infrared in vivo imaging for tumour targeting and biodistribution assays and antitumour efficacy study.
Part of Figure 3 from Zhang et al. Tumour targeting and biodistribution. (A) Representative in vivo fluorescence imaging of biodistribution at the indicated time points after i.v. injection of DiR-labelled Nano/TM (membrane of KPC cell line), DiR-labelled Nano/CD40, DiR-labelled Nano/CLDN18.2 and DiR-labelled NanoBE (nanoengager expressing anti-CD40 scFv and anti-CLDN18.2 scFv). Tumours were indicated by black circle. (B) Mean signal intensity of tumours following administration of the particles at the indicated time points. Credit: Zhang, Y., Li, Y., et.al., doi: 10.1186/s12951-024-02369-9 Reproduced under the Creative Commons license
2. Inhibition of T cell-based immunity by axon guidance molecules
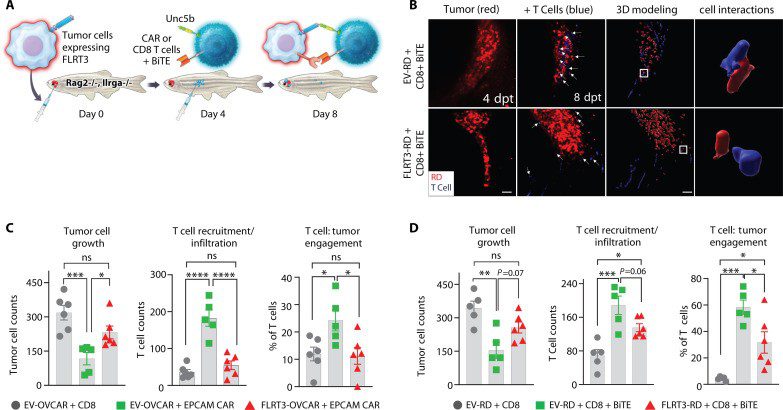
In this study, Prajapati et al. identified fibronectin leucine-rich transmembrane protein 3 (FLRT3), an axon guidance molecule (AGM), as a ligand that inhibits T cell activity. This is inhibition is through uncoordinated 5 (UNC5B), an axon guidance receptor that is upregulated on activated human T cells. Humanised zebrafish models were used to investigate the roles of the FLRT3-UNC5B axis in T cell antitumour immunity and tumour infiltration. These models were established with either ovarian cancer (OVCAR-5) or rhabdomyosarcoma (RD) cell lines and EpCAM Chimeric Antigen Receptor (CAR-T) cells or epidermal growth factor receptor (EGFR)/CD3 bispecific T cell engagers (BiTEs), respectively. The tumour cells were stably expressed with mCherry to assess tumour growth, and then intraperitoneally injected with T cells labelled with the ViaFluor 405 SE dye. The authors demonstrated that FLRT3 expression on tumours significantly reduced T cell–mediated antitumour immunity, inhibited T cell tumour environment (TME) recruitment/infiltration, and reduced T cell engagement with tumour cells. This research provides evidence that axon guidance proteins emulate T cell checkpoints, affirming their potential as targets for cancer immunotherapy.
Figure 5 from Prajapati et al. FLRT3 suppresses T cell immunity, infiltration, and tumor engagement in zebrafish models. (A) Representative schematic humanised zebrafish model. (B) Representative microscopy images showing recruitment (left two panels) and close cell interaction (engagement) (right panel) of T cells (blue) to the RD tumor cells (red). 3D modeling was used to assess T cell:tumor cell engagement. (C) OVCAR-5 cells with control CD8+ T cells, EV–OVCAR-5 + EpCAM CAR-T cells, and FLRT3–OVCAR-5 + EpCAM CAR-T cell effect on tumor growth, T cell recruitment/infiltration, and T cell engagement. (D) Same as (C) except using RD tumor model with EV-RD + EGFR-CD3 BiTEs or FLRT3-RD + EGFR-CD3 BiTEs. Credit: Prajapati, K., Yan, C., et.al., doi: 10.1126/sciadv.adj4698 Reproduced under the Creative Commons license
3. Extra centrosomes trigger mitochondrial apoptosis in DNA damage-driven tumourigenesis
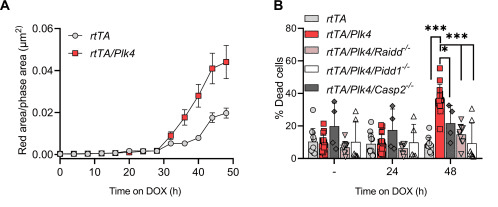
Contrary to the prevailing belief that extra centrosomes inherently promote tumor growth, Braun et. al., demonstrated that they actually induce mitochondrial apoptosis triggered by DNA damage. This process is facilitated by Polo-like kinase 4 (PLK4), which activates the PIDDosome. The study utilised NucView® 530 Caspase-3 enzyme substrate to quantify cell death in live cells. The accompanying figure illustrates that cells overexpressing Plk4 exhibited increased activation of the caspase-3 reporter, beginning approximately 30 hours after transgene induction. The data also revealed that this cell death was PIDDosome dependent; the inactivation of PIDDosome (through the loss of individual PIDDosome components – Caspase-2, Raidd, or Pidd1) could reduce the number of dead cells in the presence of Plk4 mRNA overexpression. This suggests that Plk4 overexpression engages the PIDDosome in response to extra centrosomes for the initiation of cell death. In summary, these findings reveal a new tumour-suppressive role of centrosome amplification, suggesting potential therapeutic applications in cancer treatment.
Part of Figure 6 from Bruan et al. Centrosome amplification leads to PIDDosome-dependent cell death. (A) Time-dependent caspase-3 substrate processing in multipotent hematopoietic progenitors (MPP) after doxycycline treatment was measured over time by IncuCyte analysis (red fluorescence area/phase area in μm2). (B) Quantification of DAPI and annexin V–positive MPPcells after 24 or 48 hours of doxycycline exposure. Credit: Braun, V., Karbon, G., et.al., doi: 10.1126/sciadv.adk0564 Reproduced under the Creative Commons license
4. Melanoma cells release extracellular vesicles containing lipid droplets and mitochondria during cell division
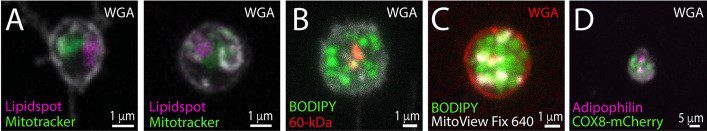
Research by Karbanova et al. revealed that melanoma cells release extracellular vesicles (EVs) containing the CD133 stem cell marker. These large EVs, observed during melanoma cell division or migration, are enriched with both lipid droplets and mitochondria. These EVs, serving as reservoirs of energy fuel, likely play a role in facilitating cancer cell spread, migration, and ultimately supporting cancer growth.
The study utilised FEMX-I cells, a highly aggressive melanoma cell line. The researchers were able to ascertain the concurrent presence of lipid droplets and mitochondria on the extracellular vesicles (EVs) by employing various experimental techniques to stain the cells. These techniques included using different combinations of fluorescent dyes to label either lipid droplets (LipidSpot™ 610, BODIPY™ 493/503) or mitochondria (MitoTracker™ Red CMXRos, MitoView™ Fix 640), as well as immunolabeling for the 60-kDa mitochondrial antigen or the lipid droplet-associated adipophilin (in cells transfected with COX8-mCherry). Cells were also co-stained with fluorescence-conjugated WGA to highlight glycoconjugates at the cell membrane.
Part of Figure 8 from Karbanova et al.FEMX-I cells were preincubated with (A) MitoTracker™ Red CMXRos or (C) MitoView™ Fix 640, and co-stained with fluorescent dyes (A) LipidSpot™ 610 or (C) BODIPY™ 493/503, or (B) immunolabelled with an anti-60-kDa mitochondrial antigen antibody and stained with BODIPY™ 493/503. (D) Alternatively, COX-8-mCherry-transfected FEMX-I cells were immunolabelled with an anti-adipophilin antibody. Samples were also co-stained with fluorescence-conjugated WGA to highlight glycoconjugates at the cell membrane. Credit: Karbanova, J., Deniz, IA., et.al., doi: 10.1186/s12964-024-01471-7 Reproduced under the Creative Commons license
5. Lipid droplet accumulation promotes proliferation and survival in prostate cancer
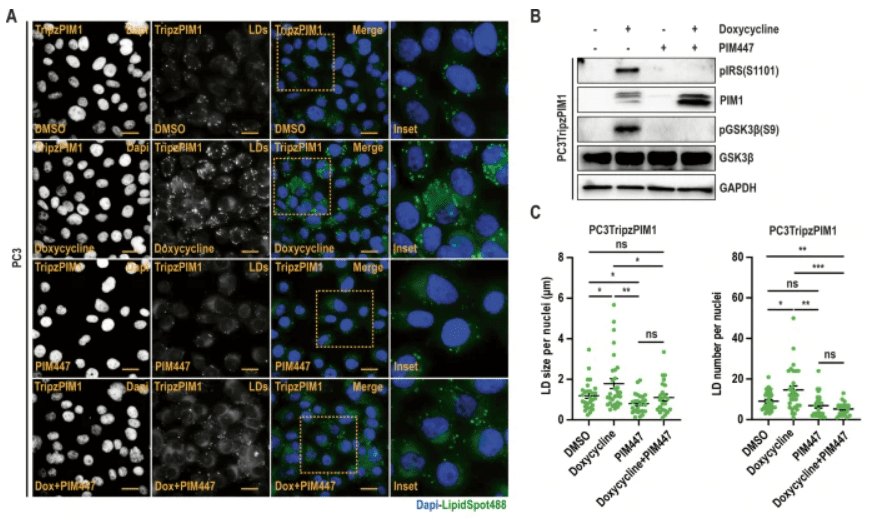
In this study, Chauhan et al. identified Proviral Integration site for Moloney murine leukemia virus 1 (PIM1) kinase as a driver of lipid droplet (LD) accumulation. They demonstrated a critical role for this event in prostate cancer cell proliferation and survival during nutrient stress. The authors showed that PIM1 acts through a novel signalling axis (GSK3β-PPARα), involving direct phosphorylation and inhibition of GSK3β, and subsequent activation of PPARα to facilitate LD accumulation. LipidSpot488 and Lipid Spot610 were utilised to assess LD accumulation in the in vitro studies.
The authors also further demonstrated that targeting PIM1 or PPARα signalling using genetic or chemical means abrogates the proliferative and survival advantage associated with LD accumulation and PIM1 induction in prostate cancer. Thus, targeting PIM1 and/or blocking LD utilisation could represent a promising strategy for prostate cancer treatment.
Part of Figure 2 from Chauhan et al. (A) Representative images of LDs in PC3TripzPIM1 cells (prostate cancer cell line stably expressing a doxycycline-inducible PIM1 vector) following doxycycline treatment alone, and in combination with PIM inhibition. (B) Western blotting of PC3Tripz-PIM1 cells following doxycycline treatment alone, and in combination with PIM inhibition. (C) Quantification of LD size and number per nuclei (n > 30 cells/group). Credit: Chauhan, S., Casilla, AL., et.al., doi: 10.1038/s41388-023-02914-0 Reproduced under the Creative Commons license
The dyes featured in these featured publications are from Biotium. A leading developer of cutting-edge stains and dyes, stands at the forefront of this revolution, providing researchers with a diverse range of high-quality reagents tailored to meet the demands of modern experimental paradigms. With a commitment to excellence and innovation, Biotium’s stains empower scientists to explore the intricacies of cellular biology with confidence and efficiency.
References
Braun VZ, Karbon G, Schuler F, Schapfl MA, Weiss JG, Petermann PY, Spierings DCJ, Tijhuis AE, Foijer F, Labi V, Villunger A. Extra centrosomes delay DNA damage-driven tumorigenesis. Sci Adv. 2024 Mar 29;10(13):eadk0564. doi: 10.1126/sciadv.adk0564. Epub 2024 Mar 29. PMID: 38552015; PMCID: PMC10980279.
Chauhan SS, Casillas AL, Vizzerra AD, Liou H, Clements AN, Flores CE, Prevost CT, Kashatus DF, Snider AJ, Snider JM, Warfel NA. PIM1 drives lipid droplet accumulation to promote proliferation and survival in prostate cancer. Oncogene. 2024 Feb;43(6):406-419. doi: 10.1038/s41388-023-02914-0. Epub 2023 Dec 14.
Karbanová J, Deniz IA, Wilsch-Bräuninger M, de Sousa Couto RA, Fargeas CA, Santos MF, Lorico A, Corbeil D. Extracellular lipidosomes containing lipid droplets and mitochondria are released during melanoma cell division. Cell Commun Signal. 2024 Jan 19;22(1):57. doi: 10.1186/s12964-024-01471-7.
Prajapati K, Yan C, Yang Q, Arbitman S, Fitzgerald DP, Sharee S, Shaik J, Bosiacki J, Myers K, Paucarmayta A, Johnson DM, O’Neill T, Kundu S, Cusumano Z, Langermann S, Langenau DM, Patel S, Flies DB. The FLRT3-UNC5B checkpoint pathway inhibits T cell-based cancer immunotherapies. Sci Adv. 2024 Mar;10(9):eadj4698. doi: 10.1126/sciadv.adj4698. Epub 2024 Mar 1.
Zhang H, Li Y, Kang H, Lan J, Hou L, Chen Z, Li F, Liu Y, Zhao J, Li N, Wan Y, Zhu Y, Zhao Z, Zhang H, Zhuang J, Huang X. Genetically engineered membrane-based nanoengagers for immunotherapy of pancreatic cancer. J Nanobiotechnology. 2024 Mar 11;22(1):104. doi: 10.1186/s12951-024-02369-9.
Microscopes have been our eyes into the invisible world, revealing intricate details of life and matter. From the tiniest microorganisms to the intricate structures of
Summary Get an insightful overview of the burgeoning field of cell and gene therapies, showcasing their potential to revolutionize medical treatments. Discover the power of
Discover current clinical trials using Mesenchymal Stem Cells (MSCs) for cancer therapy. Learn how MSCs are being studied to treat cancer and alleviate side effects of traditional therapies.
HOW CAN WE HELP YOU?Our specialists are to help you find the best product for your application. We will be happy to help you find the right product for the job.